Chapter 18: Smith and Nathans discover enzymes that dissect DNA
“… the restriction endonuclease should prove of general usefulness in the analysis of DNA, much as highly specific proteolytic enzymes have been used in the analysis of proteins.”
The genetic material in chromosomes is a single, uninterrupted DNA double helix. How, then, did it become possible for molecular biologists to isolate and study individual genes and their regulatory elements? The breakthrough that helped make this possible and helped launch a new era in molecular biology was the discovery of endonuclease restriction enzymes. This is a story that played out over 20 years in three stages.
The discovery of restriction modification. The story begins in the early 1950s with our hero Salvador Luria (chapter 2), who was studying the infection of E. coli by phage T2 (a very close relative of the phage T4 in chapters 9 and 10). He observed that a mutant of E. coli was killed by T2 phage but seemed not to release viable phage progeny that could infect wild-type E. coli. As Luria recounts in his autobiography, A Slot Machine, A Broken Test Tube, “One day the test tube containing the phage-sensitive bacterial culture happened to break.” Rather than give up on his experiment, he obtained from a colleague a sample of a different kind of bacterium called Shigella. Lo and behold, the mutant E. coli had released viable phage, but these phage were modified in some way such that they could grow on Shigella but no longer on E. coli. Luria had obtained the first evidence for the phenomenon known as restriction modification.
As we now know, bacteria have defense mechanisms that protect them from phage and certain phage have counter defense systems. Phage T2 evolved a modified base, hydroxymethyl cytosine, to protect it from restriction enzymes. But as a counter defense strategy, E. coli evolved a restriction enzyme that recognizes and cleaves hydroxymethyl-cytosine-containing DNA. Not to be outdone, T2 further evolved to modify its DNA by attaching a protective sugar (glucose) residue to its hydroxymethyl cytosines. A phage enzyme transfers glucose to hydroxymethyl cytosine from an activated form of glucose called UDP-glucose. The resulting “sweet” DNA is protected from the host restriction enzyme. We now know that Luria’s mutant E. coli lacked the UDP-glucose precursor needed for sugar transfer to the hydroxymethyl cytosines. Hence the progeny phage were “sour” and could no longer grow on E. coli. Shigella, however, lacks the sour DNA-degrading enzyme and can therefore support the growth of sweet or sour DNA. Thus, when Luria used Shigella, the progeny phage that had been grown on the mutant E. coli were fully capable of propagation on the alternative host.
Luria’s “broken test tube” experiment is more complicated than restriction modification systems that don’t have counter defense systems. Nonetheless, it is historically significant because it was the first clue that led to the discovery of endonuclease restriction enzymes. After Luria’s findings, numerous other examples of restriction modification systems were discovered. But the first demonstration that restriction acted at the level of DNA degradation was made in the 1960s by Swiss microbiologist and geneticist Werner Arber (1929-) using phage λ. Arber had learned phage genetics from Joe Bertani at the University of Southern California and had visited the labs of Luria and of Esther and Joshua Lederberg. His subsequent work at the University of Geneva showed that DNA from phage that had been grown on E. coli strain K was degraded shortly after infection of E. coli strain B.
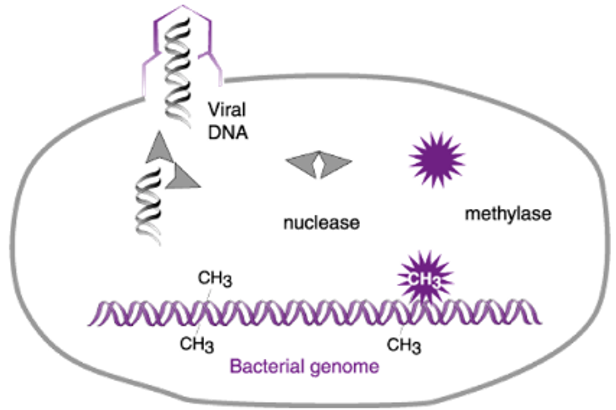
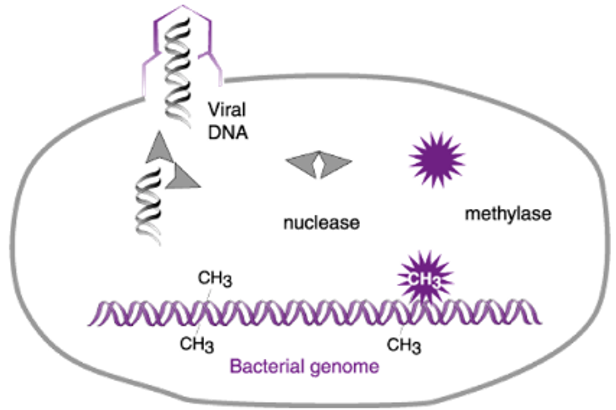
We now know that E. coli strain B produces both a restriction enzyme (nuclease in the cartoon) and a modification enzyme (methylase). The restriction enzyme recognizes a short sequence prevalent in both host and phage DNAs and protects its own DNA from cleavage by attaching methyl groups to certain nucleobases in this sequence. Phage λ that was grown on strain K lacks these protective methyl groups and hence is destroyed upon infection of strain B.
Discovery of endonuclease restriction enzymes. This discovery was reported by Linn and Arber in 1968. A key to their finding was that the enzyme required S-adenosyl methionine and ATP as co-factors. A few months later a similar discovery was reported by Yuan and Meselson. The Linn and Arber paper proved to be controversial in that the authors did not make it clear that they had evidently learned of the co-factor requirements from Yuan and Meselson, merely stating that the requirement “confirms a finding made in another system by Yuan and Meselson which had come to our attention during our experiments.” Since Arber would share in the Nobel Prize that was awarded for the discovery of restriction endonucleases (as we will come to) and Meselson not, the controversy was not of little significance. Meselson explains in the video how he and Yuan came to discover the requirement for S-adenosyl methionine and ATP and that they shared this information widely prior to publication.
Two final points about the kind of endonuclease restriction enzymes discovered by Linn and Arber and by Yuan and Meselson are important. First, these enzymes are a three-protein complex that is responsible both for methylating host DNA at the recognition sequence and for cleavage of unmodified DNA. Second, and although the enzymes act by recognizing a specific sequence in DNA, they don’t cleave the DNA at the recognition sequence. Rather, they cleave DNA at a random distance from the recognition sequence. Thus, although the discoveries of the first endonuclease restriction enzymes were of historical importance, the enzymes themselves were not useful for cutting DNA at unique sites.
The discovery of an endonuclease restriction enzyme that cleaves DNA at the recognition sequence. The heroes of the final part of the endonuclease restriction enzyme story are Hamilton Smith and Dan Nathans. Hamilton “Ham” Smith (1931-2025) received his MD from Johns Hopkins School of Medicine in 1956 and did research at Washington University, the University of Zürich and the University of Michigan, where he read the article in “…the Journal of Molecular Biology containing the classical Jacob and Monod paper describing the operon model for gene regulation, and …..I suddenly became aware of the beautiful work of the “phage school” and of Watson and Crick and DNA. After many years of haphazard searching for the “right” area of research, I knew I had found it.” He then joined the faculty of Johns Hopkins University School of Medicine as an assistant professor of microbiology in 1967, later becoming deeply involved in the emerging field of genomics. Dan Nathans (1928-1999) received his MD from Washington University in 1954. He then did research with Nobel Laureate and biochemist Fritz Lippman at Rockefeller University and joined the faculty of Johns Hopkins University School of Medicine in 1962. He would later serve as president of the University. Smith and Nathans won the Nobel Prize with Werner Arber in 1978.
In two back-to-back publications, Smith reported the discovery of a restriction enzyme (now called HindII) from the bacterium Hemophilus influenzae that cleaved DNA at specific sites, writing that the enzyme was able to “recognize only a few specific sites on rather large foreign DNA molecules.” From the size of the fragments it generated using various phage DNAs as substrates, he was able to estimate that the recognition sequence “would have to be five or six bases in length…” In the second of the two publications, he reported the sequence of this recognition site, which exhibited a two-fold rotational axis of symmetry (a palindrome) and was indeed six base pairs:
5’…GTYRAC…3’
3’…CARYTG…5’
where Y is either a pyrimidine (C or T) and R a purine (A or G). He further showed that cleavage at the recognition sequence results in blunt ends (that is, there are no protruding 5’ or 3’ single-stranded ends):
5’…GTY RAC…3’
3’…CAR YTG…5’
In chapter 19, we will come to an endonuclease restriction enzyme called EcoR1 that is more restrictive than Smith’s Hemophilus enzyme. EcoR1 was discovered in the laboratory of Herb Boyer at the University of California, San Francisco in 1971. As in the case of the Hemophilus enzyme, the recognition site for EcoR1 exhibits a two-fold axis of symmetry and is six-base pairs in length. But the unlike the Hemophilus recognition sequence, the EcoR1 recognition sequence requires a specific purine or pyrimidine at each position:
5’…GAATTC…3’
3’…CTTAAG…5’
Also, unlike the Hemophilus enzyme, which generates blunt ends, EcoR1 makes two-single stranded, staggered cleavages to create two four-nucleotide-long, complementary 5’ overhangs:
5’-AATTC…3’
G…5’
5’…G
3’…CTTAA-5’
EcoR1 became famous with the advent of gene cloning, which it facilitated, as we shall see, by virtue of the specificity of its recognition sequence and the 5’ overhangs (“sticky ends”) it generates.
Finally, it was Smith’s colleague, Dan Nathans, who demonstrated how an endonuclease restriction enzymes could be used to dissect DNA. In his 1971 publication, Nathans reported that Smith’s Hemophilus influenzae restriction enzyme could cleave a cancer-causing virus, Simian Virus 40 (SV40), into eleven fragments, which he could visualize by gel electrophoresis (lower image). This enabled him to conclude that, “The availability of pieces of SV40 DNA from specific sites in the molecule should be helpful for the analysis of the function of the SV40 genome. ….Therefore, the restriction endonuclease should prove of general usefulness in the analysis of DNA, much as highly specific proteolytic enzymes have been used in the analysis of proteins.” This, indeed, proved to be the case.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}