Chapter 17: Zamecnik discovers the tRNA adaptor
“It is therefore a natural hypothesis that the amino acid is carried to the template by an adaptor molecule, and that the adaptor is the part which actually fits on to the RNA.”
Among the prescient hypotheses advanced by Crick in “On Protein Synthesis” (Chapter 14) was the Adaptor Hypothesis (as enunciated in the quote above). The discovery that genetic information is transported to the ribosome by messenger RNA demanded the existence of an adaptor between the nucleotide codons and the machinery for stringing amino acids together into protein. Indeed, Crick imagined an adaptor for each amino acid. The pioneering biochemist who discovered these adaptors was Paul Zamecnik (1912-2009).
Zamecnik obtained his Medical Degree from Harvard Medical School in 1936. He did research in Boston, Copenhagen and Rockefeller before returning to Harvard Medical School as an Assistant Professor in 1942. He did his classic work on tRNA at the Huntington Memorial Hospital of Harvard (since demolished) and then Massachusetts General Hospital. After retiring in 1979 he moved to the Worcester Foundation, which later merged with the University of Massachusetts Medical School, where he developed the use of “anti-sense” RNAs as a therapeutic. Zamecnick won the national Medal of Honor in 1991 and the Lasker Award in 1996.Zamecnick’s long-time collaborator Mahlon Hoagland (1921-2009) obtained a Medical Degree from Harvard Medical School in 1948. A bout with tuberculosis derailed his plans to become a pediatric surgeon. Instead, he entered the laboratory of Zamecnik to do research. He went on to become an Associate Professor at Harvard Medical School from 1953 to 1967 and Chair of Biochemistry at Dartmouth Medical School in 1967. Hoagland spent a year working with Crick in Cambridge and moved to Worcester Foundation in 1970, where he was later joined by Zamecnik.
Zamecinik, Hoagland and coworkers made the seminal discovery that amino acids are transferred to an RNA adaptor as an intermediate in protein synthesis. This RNA was successively called “pH 5 RNA” (based on a precipitation step described below), “sRNA” (for soluble RNA) and, finally, “tRNA” (for transfer RNA), the name now in use.
Hoagland et al. summarized the process by which amino acids are transferred to RNA as a two-step process. In the first step, (reaction 1), an enzyme (E, now known as amino acyl tRNA synthetase) generates an activated form of an amino acid (leucine in the example) by reaction with ATP. The amino acid in this “amino acyl-adenylate” is linked to AMP by a high energy bond (as represented by the ~ symbol). In the second step the enzyme transfers the activated amino acid to an RNA, releasing AMP. We now refer to the RNA as a “charged tRNA”.
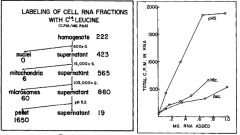
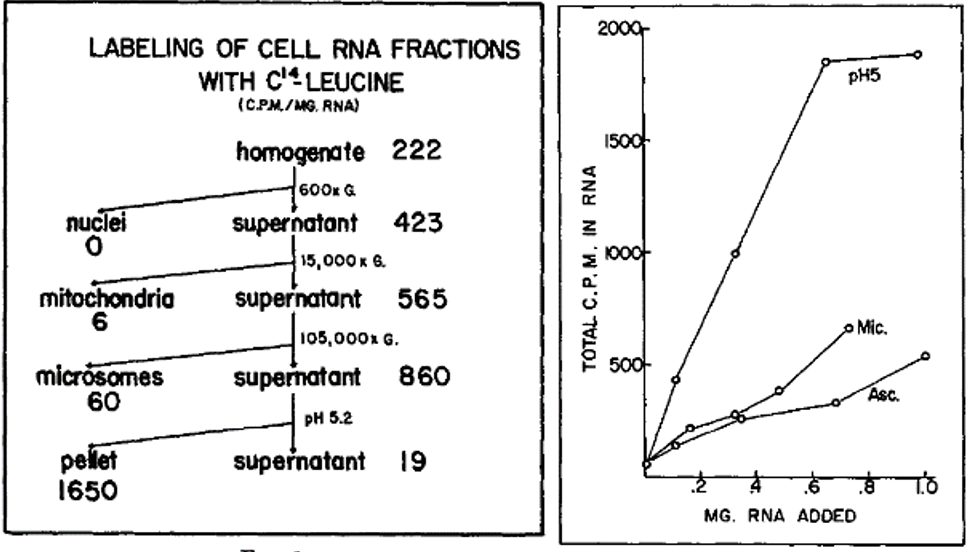
The left-hand figure summarizes the purification steps that led to the discovery of the enzyme and the RNA. Note that the pellet from a precipitation step (the pellet) carried out at pH 5 was enriched in an activity that would incorporate radioactive leucine (leucine-C14) into RNA. As an example of their findings, the right-hand figure shows that RNA extracted from the pH 5 precipitation step was more effective in stimulating the incorporation of radioactive leucine into RNA than was RNA extracted from microsomes (Mic), a ribosome containing fraction, or from ascites cells (Asc).
In the video below Zamecnik recounts some of the steps in the discovery tRNA charging:
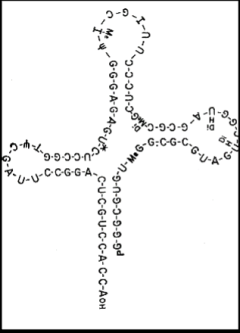
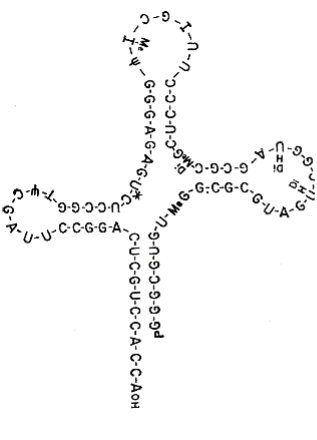
The next major advance in the story was made by Robert Holley (1922-1993). Holley obtained his PhD at Cornell University in Organic Chemistry in 1947 and joined the faculty at Cornell in organic chemistry the following year. He spent a year at CalTech in 1955-56 studying RNA. Returning to Cornell he pioneered the sequencing of RNA (beating Sanger who also attempted to sequence RNA; Chapter 11), focusing on tRNA and alanyl tRNA in particular. He shared the Nobel Prize in 1968 with Nirenberg and Khorana. The images below are from his classic 1965 publication in Science and include the sequence of alanine tRNA. Note that tRNA contains modified bases, such as dimethyl guanine (DiMe) and pseudouridine (ψ) as indicated in the figure.
Deserving of special mention is Elizabeth Keller, who had been part of Zamecnik’s team at Harvard and later moved to Cornell. She devised the cloverleaf model for the secondary structure of tRNA. In this ionic model, the site of amino acylation at which an amino acid is attached is at the 3’ end of the RNA at the bottom of the stem of the cloverleaf and the anti-codon is at the opposite end of the structure in the top loop (leaf). According to her obituary in the New York Times (below), she sent her model to Holley in a Christmas card. He included the model in his Nobel address but did not acknowledge her (although he did share some of the Prize money with her and others in his team).
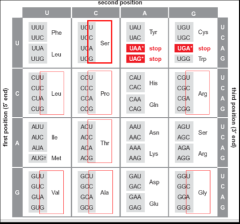
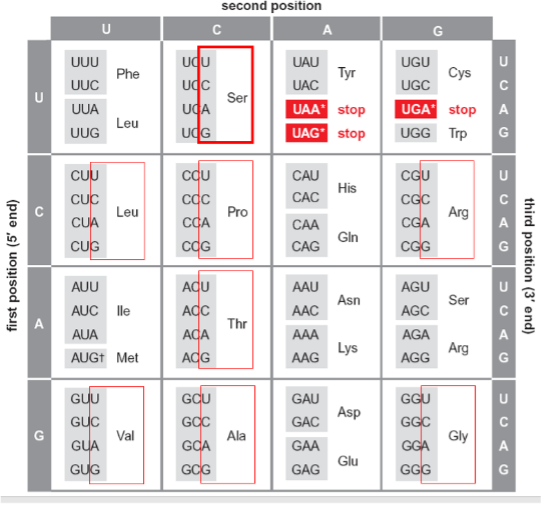
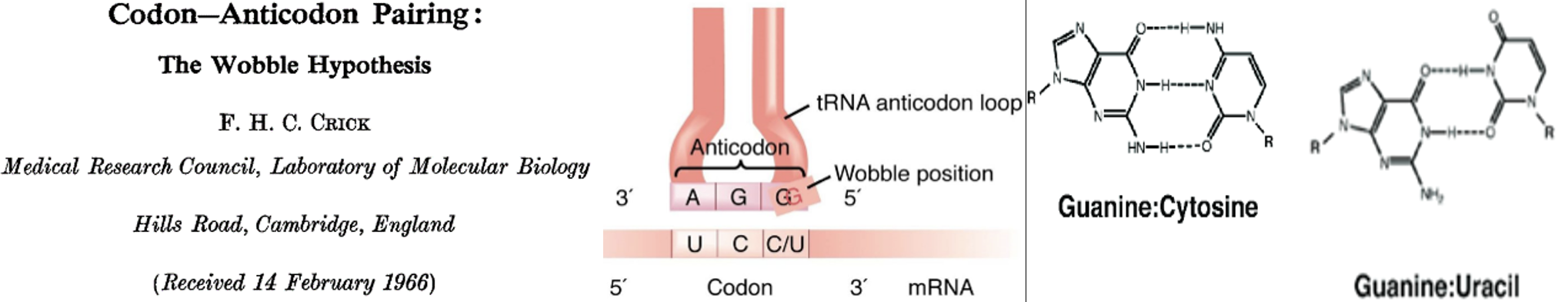
Ever the prescient theoretician, Crick made one more impactful contribution to the adaptor story following on the cloverleaf structure and the identification of the anti-codon. As Crick, Barnett, Brenner and Watts-Tobin had pointed out in 1961, a three-letter code for twenty amino acids demanded degeneracy. Indeed, all of the 20 amino acids save methionine and tryptophan are specified by more than one codon. This degeneracy is manifest, wholly or in part, in the 3’ position of the codon. Thus, as highlighted in dark red, serine is specified by four codons that differ simply at the 3’ position. The same is true for alanine, arginine, glycine, leucine, proline, and valine as highlighted in red. In addition, alternative bases at both the first position and at the third allow recognition of the same amino acid by six codons, as is the case for serine and arginine. Crick hypothesized that “wobble” between the pairing of the third base (3’) and the first (5’) base of the anti-codon in the tRNA could allow the same tRNA to recognize more than one codon for the same amino acid. Thus, as illustrated in the cartoon (middle), wobble of the G at the 5’ position of the AGG anticodon would allow recognition of both 5’ UCC 3’ and 5’ UCU 3’ as serine codons. For example, guanine can pair with either cytosine or uracil (right) when their interaction is not as tightly constrained as in an extended double helix. We now know that the number of tRNAs isoacceptors varies among species but all known organisms employ wobble to recognize synonymous codons at least to some extent.
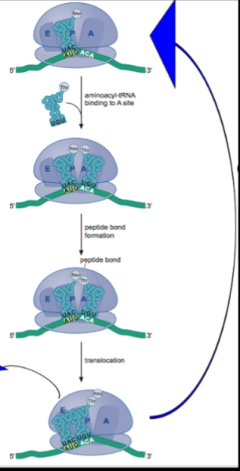
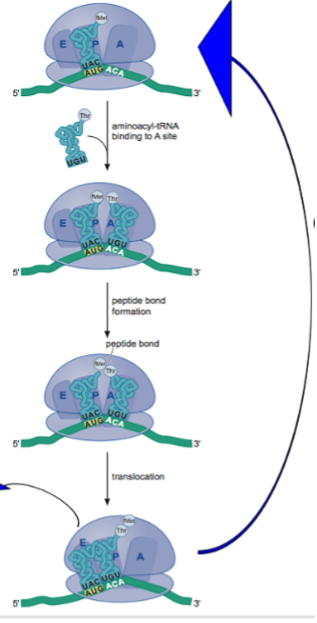
In the ensuing years much progress was made in elucidating fundamental features of the machinery for translating mRNA into protein, history that is beyond the scope of this eBook. Here, we simply summarize the principal features of our understanding of a cycle of peptide bond formation that emerged from this work. First, each tRNA is charged with the correct amino acid by a dedicated tRNA synthetase. Next, as shown in the cartoon, an amino acyl tRNA (“charged” tRNA) with the correct codon/anticodon complementarity is delivered to the A site of the ribosome. A peptide bond then forms between the amino acid (methionine in the cartoon or a growing peptide chain) that is attached to the 3’ end of the tRNA in the P site. This results in transfer of the methionine (or a growing chain) to the amino acyl amino acid (threonine in the cartoon) in the A site. Finally, the ribosome translocates one codon unit along the mRNA, moving the tRNA that had been in the P site and is no longer charged to the E site where it is released from the ribosome. Meanwhile, because of translocation the A site is now vacant and can accept a new charged tRNA in the next cycle of peptide bond formation. Not shown is that an “initiation” factor delivers the correct charged tRNA (that is, having the correct anticodon) to the A site, in a manner that involves the hydrolysis of a molecule of GTP and that another protein factor, an “elongation” factor, which also uses GTP as an energy source, is responsible for the translocation of the ribosome along the mRNA.
All this was captured artistically and humorously in a famous 1971 dance video introduced by Paul Berg of Stanford University, who would later win a Nobel Prize for the invention of recombinant DNA (chapter 19). In the video, reflecting the hippy culture of the day, the participants dynamically act out the steps in protein synthesis outlined above. Note that the delivery of a charged tRNA to the ribosome by a protein factor is accompanied by a puff of smoke representing GTP hydrolysis and that translocation of the ribosome by the elongation factor is also accompanied by a puff of smoke from GTP hydrolysis.
This page has paths:
This page references:
- Crick Codon Degeneracy
- Crick Codon Wobble
- Hoagland RNA Transfer
- Holley Portrait
- Keller Obituary
- Keller tRNA Model
- mRNA Protein Machinery
- RNA Enzyme Discovery
- Zamecnik Hoagland Intermediate
- Zamecnik Portrait
- Chapter17RibonucleicStructureDiagram
- Chapter17StructureOfRibonucleicAcidHeader
- Chapter17HolleryEverettApgarMadison
{kind=link}
{kind=link}
{kind=link}
{kind=link}
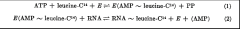{kind=link}
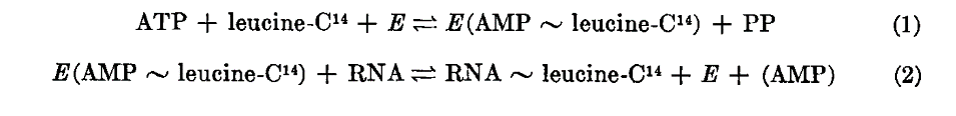{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
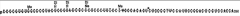{kind=link}
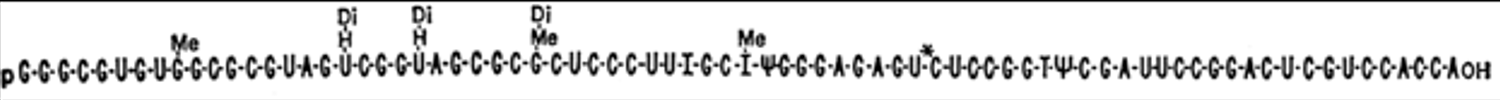{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}