Chapter 14: Crick Enunciates the Central Dogma and Two Research Teams Discover Messenger RNA
“…once ‘information’ has passed into protein it cannot get out again.”
In his highly impactful treatise “On Protein Synthesis” published in 1958, Francis Crick spelled out two foundational concepts of molecular biology, The Sequence Hypothesis and The Central Dogma. The former holds that “the specificity of a piece of nucleic acid is expressed solely by the sequence of its bases, and that this sequence is a (simple) code for the amino acid sequence of a particular protein.” Indeed, this foreshadowed his epic publication with Barnett, Brenner and Watts-Tobin in 1961. The second concept states that “once ‘information’ has passed into protein it cannot get out again. In more detail, the transfer of information from nucleic acid to nucleic acid, or from nucleic acid to protein may be possible, but transfer from protein to protein, or from protein to nucleic acid is impossible. Information means here the precise determination of sequence, either of bases in the nucleic acid or of amino acid residues in the protein.” These two concepts posed the question of how information in the sequence of bases is converted into a sequence of amino acids.
The earliest suggestion that information in the sequence of the gene is transmitted via an unstable RNA intermediate was made, albeit hesitatingly, in 1954 by Jacob and Monod’s collaborator in the PaJaMo experiment, Art Pardee. He wrote, “One could visualize an active form of RNA…..which, once used to mold a specific protein molecule, becomes an inert waste product [underlining added] in E. coli.” Based on follow up experiments in which he measured the kinetics of enzyme synthesis after mating in 1958, Pardee more explicitly stated, “A second class of hypotheses assumes that DNA does make an intermediate carrier of information (perhaps RNA). One must then explain why the rate of formation of enzyme remains constant per cell after mating; and a number of possible but not too plausible explanations exist, based on the idea that the cell is saturated with templates in a few minutes, or that the number of templates reaches equilibrium in a few minutes, or that the templates might be unstable and rapidly reach a steady-state concentration [underlining added] as a balance of formation and inactivation. This hypothesis of templates with short lifetimes has been suggested earlier on the basis of the continued requirement of components of nucleic acids for protein synthesis [his 1954 publication].
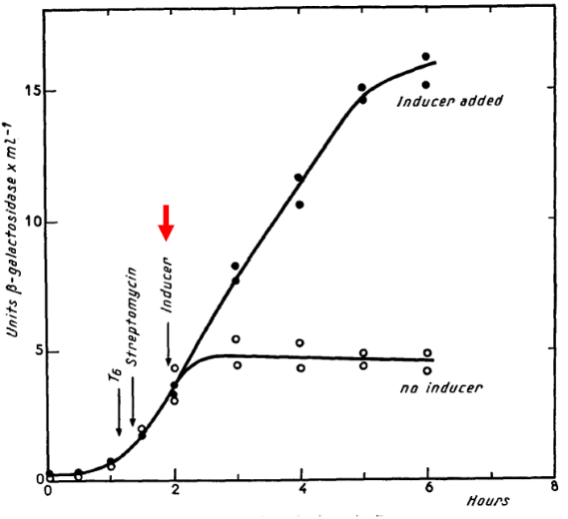
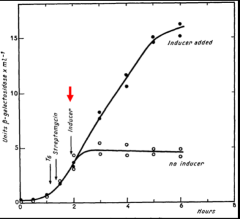
The unstable messenger RNA hypothesis is most explicitly stated by Jacob and Monod in 1961, “The property attributed to the structural messenger of being an unstable intermediate is one of the most specific and novel implications of this scheme; it is required, let us recall, by the kinetics of induction, once the assumption is made that the control systems operate at the genetic level. This leads to a new concept of the mechanism of information transfer, where the protein synthesizing centers (ribosomes) play the role of non-specific constituents which can synthesize different proteins, according to specific instructions which they receive from the genes through M-RNA.” Notice in the figure from their mating experiment that β-galactosidase synthesis rapidly reaches steady state when inducer (highlighted with red arrow) is added but almost immediately ceases when inducer is absent. Oddly, Jacob and Monod failed to cite the 1954 and 1958 publications by Pardee in their landmark 1961 publication. Perhaps, Pardee had simply been too cautious.
The wider community was slow to accept the concept of an unstable messenger. In his autobiography, Jacob recounts the reaction to a presentation he gave in Copenhagen in 1959 in which he stressed the need for an unstable intermediate, “No one reacted. No one batted an eyelash. No one asked a question”.
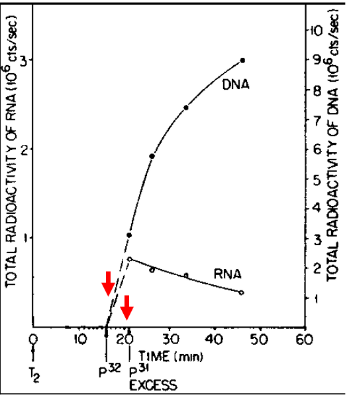
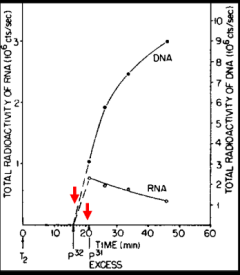
Meanwhile, an important, but slow to be appreciated clue was the discovery by Elliot Volkin and Lazarus Astrachan at the Oak Ridge National Laboratory in 1957 of unstable RNA in phage-infected cells. They carried out a “pulse-chase” experiment in which cells that had been infected with a phage were briefly radioactively labeled with P32 followed shortly thereafter by exposure to an excess of non-radioactive phosphorous (P31). As highlighted by the red arrows in the figure, radioactivity was incorporated into an unable RNA species that decreased in abundance during the chase. The authors and others missed the significance of this unstable RNA. It took until 1960 at an informal gathering at which Jacob, Brenner and Crick were present for the meaning of the Volkin and Astrachan experiment to suddenly become appreciated. As recounted by Jacob, “At this precise point, Francis and Sydney leaped to their feet. Began to gesticulate. To argue at top speed in great agitation. A red-faced Francis. A Sydney with bristling eyebrows. The two talked at once, all but shouting. Each trying to anticipate the other. To explain what had suddenly come to mind…What had until then been an abstraction was becoming a molecular species... this RNA of the phage was indeed the unstable intermediary functioning in the synthesis of proteins.” Voilà!
The pièces de résistance in the discovery of messenger RNA were two back-to-back publications in Nature in 1961 that provided experimental evidence for an unstable intermediate in the transfer of genetic information to the site of protein synthesis, ribosomes.
It was generally understood that proteins could not be synthesized directly on genes; consider, for example, eukaryotic cells in which chromosomes are segregated from the machinery for protein synthesis, ribosomes, which are located in the cytoplasm. Importantly, these publications distinguished between two competing models. In one model, ribosomes (also referred to as “RNA particles” before their function was fully understood) are each dedicated to the synthesis of a particular protein through their RNA component. In the other model, ribosomes are non-specialized factories that can synthesize at any given time any particular protein as dictated by the messenger RNA with which they are associated.
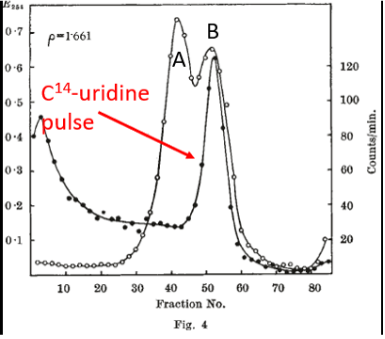
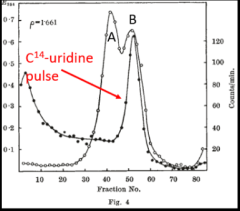
At the heart of the Brenner, Jacob and Meselson report was a pulse-labeling experiment carried out after phage infection, which was designed to ask whether newly synthesized RNA became associated with ribosomes. The infected cells were pulse-labeled with radioactive (C14) uridine (Fig. 4 on the left) after phage infection. After labeling, centrifugation was carried out to separate macromolecules according to density. Note that ribosomes consist of two subunits. Intact ribosomes (open circles), which are labeled B in Fig. 4, sedimented at about fraction 50. [Ribosomes that had dissociated into their subunits (labeled A in Figure 4) sedimented more rapidly (fractions to the left.] The key result was that newly synthesized RNA (filled circles in Fig. 4) associated with intact ribosomes. Following the pulse of C14-uridine, infected cells were chased with an excess of non-radioactive (C12) uridine (Fig. 5 on the right). It can be seen that the peak of newly synthesized RNA associated with intact ribosomes diminished substantially following the chase, a finding consistent with the idea that the ribosome-associated, pulse-labeled RNA was unstable. In toto, these results provided support for the existence of an unstable messenger RNA that became associated with ribosomes following phage infection.
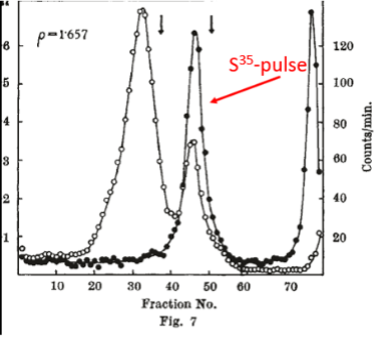
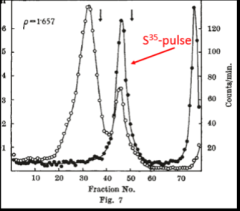
Next, Brenner, Jacob and Meselson went a step further. To distinguish pre-existing ribosomes from ribosomes synthesized after phage infection, cells of E. coli were grown on “heavy” medium containing the heavy (but not radioactive) isotopes nitrogen-15 and carbon-13 (see historical footnote below) and phosphorous-32 as a radioactive label for ribosomes. The bacteria were then infected with phage and shifted to “light” medium containing nitrogen-14 and carbon-12 and pulse-labeled with S35-suphate to label protein synthesized after phage infection (highlighted in red in Fig. 7). Finally, after the pulse-labeling, centrifugation was carried out to separate macromolecules according to density. The key result was that newly synthesized protein (filled circles in Fig. 7) was associated with heavy ribosomes. (The expected positions of light, A and B ribosomes is indicated by the two vertical arrows.) The authors concluded that protein in infected cells is synthesized on pre-existing ribosomes. To quote from the authors, “…the experiments with phage-infected cells show unequivocally that information for protein synthesis cannot be encoded in the chemical sequence of the ribosomal RNA. Ribosomes are non-specialized structures which synthesize, at a given time, the protein dictated by the messenger they happen to contain.” Similar experiments carried out by Gros et al. showed that pulse-labeled RNA also associates with intact ribosomes in uninfected cells. Again, to quote from the authors, “Our working hypothesis is that no fundamental difference exists between protein synthesis in phage-infected and uninfected bacteria. In both cases typical ribosomal RNA does not carry genetic information, but has another function, perhaps to provide a stable surface on which transfer RNA's can bring their specific amino-acids to the messenger RNA template.”
Finally, we note that it was no accident that the Brenner et al. and Gros et al. manuscripts were published back-to-back in Nature. Watson, the senior author on the Gros et al. publication, had famously sent a telegram to Brenner asking him to hold up (“delay”) his manuscript so that both teams could publish simultaneously. Indeed, Brenner did.
An historical footnote. We end this chapter with the story of the source of the highly enriched carbon-13. Nitrogen-15 was readily available in the United States but not highly purified carbon-13. But Meselson wanted both heavy isotopes to maximize separation of pre-existing ribosomes from newly synthesized ribosomes by density gradient centrifugation. Meselson’s mentor, Linus Pauling (Chapter 6), had recently (1958) been elected to the Soviet Academy of Sciences. Pauling asked his colleagues in the Soviet Union if they would purify carbon-13 for Meselson. It took a year and required a two-story diffusion apparatus. But eventually they produced highly purified carbon-13, which was sent to Meselson. Meselson then fed the carbon-13 in the form of carbon dioxide to algae. The algae were then hydrolyzed and included in the medium for the growth of the E. coli cells. Thus, the historic Brenner, Jacob and Meselson experiment was facilitated by an extraordinarily generous gift of a rare isotope that required an elaborate and time-consuming purification process carried out on behalf of the renowned chemist Linus Pauling! Oddly, none of this is mentioned in the Nature publication except that carbon-13 was fed to E. coli in the form of an algal hydrolysate. Below are excerpts from Pauling’s 1958 letter to A. N. Nesmeyanov, President of the Academy of Sciences of the Union of the Soviet Socialist Republics:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
This page has paths:
This page references:
- Astrachan Volkin RNA DNA Radioactivity
- Astrachan Volkin RNA Turnover
- Brenne Jacob Unstable intermediate 1
- Brenner Jacob C14 Pulse Graph
- Brenner Jacob S35 Pulse Graph
- Brenner Jacob Unstable Intermediate 2
- Crick Protein Synthesis
- PaJoMo ZI Mutation Graph
- Linus Pauling Carbon Sourcing Letter
- Chapter14WatsonBrennerTelegramV2
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}