Sanger DNA Polymerase Sequencing
1 2025-01-22T09:20:34-05:00 George Shaohua Qiao 78e5371050dd5c0e21d36bad469c70d8d4be5464 225 1 Sanger's 1975 report on a new sequencing method plain 2025-01-22T09:20:34-05:00 George Shaohua Qiao 78e5371050dd5c0e21d36bad469c70d8d4be5464This page is referenced by:
-
1
2025-01-22T09:20:25-05:00
Chapter 20: Gilbert and Sanger Invent Methods for Sequencing DNA
4
plain
2025-04-03T11:21:02-04:00
“A simple and rapid method for determining nucleotide sequences in single-stranded DNA by primed synthesis with DNA polymerase is described.”
The next technological advance that revolutionized our understanding of genes and genomes and of evolution was the invention of methods to determine the nucleotide sequence of DNA. This story begins with two giants whom we have encountered in earlier chapters for their contributions to molecular biology. These individuals are Walter Gilbert (chapters 13 and 14), who pioneered the isolation of a regulatory protein and who helped discover mRNA, and Fred Sanger (chaper 11), who determined the first sequence of an entire protein.
Gilbert’s invention of DNA sequencing begins with his interest in the operator binding site for the lactose operon repressor LacI. Gilbert and his student Allan Maxam were able to isolate the operator by binding LacI to the operator and then using a deoxyribonuclease (DNase) to digest flanking DNA that was not in contact with the repressor. To obtain its sequence, they employed a painstaking (“it took us two years,” according to Gilbert) procedure in which they generated RNA copies of the DNase-protected DNA, cleaved the RNA into fragments, and then used previously developed methods for sequencing the RNA fragments.
Importantly, in 1975, Russian molecular biologist Andrei Mirzabekov visited Gilbert at Harvard. He had developed a method based on the chemical dimethyl sulfate for selectively methylating guanine and adenine bases in DNA, preferentially methylating the former over the latter. Gilbert recounts his visit and its impact in the video.
This led to a collaboration. Gilbert, Maxam and Mirzabekov showed that methylation of purines destabilized the glycosidic bond that joins purines to the deoxyribose sugar, resulting in release of the purine. Subsequent treatment with alkali could then be used to selectively break the bond between the purine-free sugar and the backbone phosphate, resulting in a break in the DNA. Importantly, limiting levels of the chemical treatments were used, so that cleavage occurred at only a fraction of the guanosine and adenosine nucleotides at any given position. Hence, when the fragmented DNA was subjected to gel electrophoresis a ladder of fragments were observed, representing the positions of guanine and adenine in the DNA.
As reported in their 1976 publication, Gilbert, Maxam and Mirzabekov applied this procedure to operator DNA that was radioactively labeled at one end with P32-phosphate. Methylation with dimethyl sulfate followed by alkali treatment generated a ladder of end-labeled fragments, giving a stronger signal (more breakage) at guanine nucleotides than at adenine nucleotides. By comparing the pattern of fragments between repressor-bound and unbound DNAs, they could identify positions of close contact between LacI and its operator, that is, positions at which repressor binding involved contact with a guanine or adenine residue would not be methylated. Indeed, since the site of methylation of guanine (N7) is in the major groove and that for adenine (N3) in the minor groove, they could distinguish sites of contact between the two grooves of DNA.
Gilbert subsequently realized that if additional chemical treatments could be devised that distinguished among all four bases, a general method could be developed for sequencing DNA. Indeed, and together with Allan Maxam, three additional treatments were devised for that preferentially cleaved at adenine over guanine, equally cleaved at cytosine and thymine, and selectively cleaved at cytosine, as reported in their historic 1977 publication.
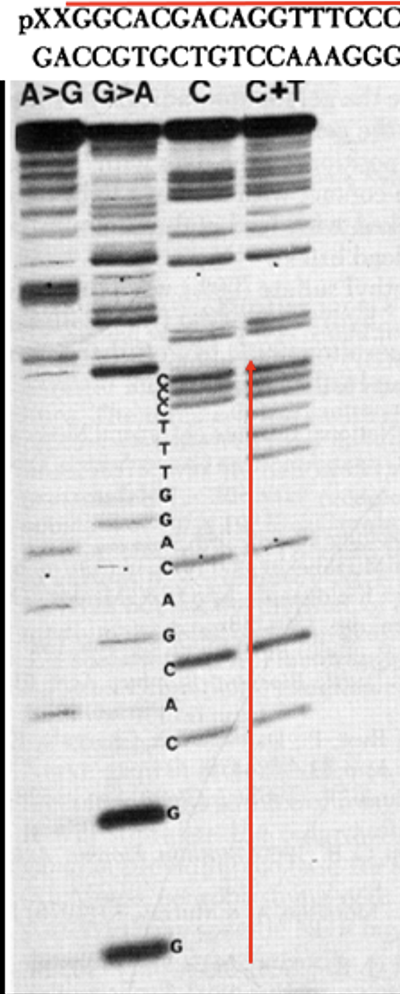
As an example, the figures from their publication show an inferred sequence (top) and the corresponding experimental sequencing ladder (bottom) upon which the inferred sequence is based. Red line and red arrow have been added to link the ladder to the deduced sequence. Thus, it can be seen that the four reactions yielded a sequence of 5’-GGCAC… (5’ because the radioactive phosphate is at the 5’ end and the smallest fragments with the 5’ label migrated the furthest down the gel). Researchers in the Gilbert laboratory then applied the Maxam Gilbert procedure to sequence the gene for the lactose repressor LacI and an entire 4,362 nucleotide pair-long E. coli plasmid. Maxam Gilbert sequencing came to be widely adapted by other researchers in the late 1970s.
Sanger created a different method for sequencing DNA and did so in two iterations. In the first, he used the DNA to be sequenced as a template for DNA synthesis. Importantly, he used limiting concentrations of each of the four deoxynucleotide substrates in separate reactions. This generated a ladder of fragments that could be resolved and visualized by gel electrophoresis. As in Maxam/Gilbert sequencing the ladder could be used to deduce the sequence of the template DNA.
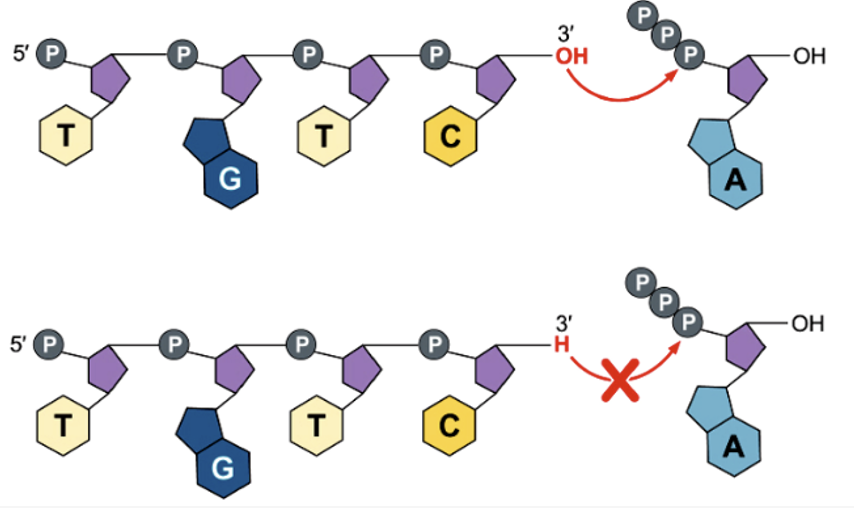
Two years later Sanger reported a striking improvement in his sequencing methodology based on the use of 2’, 3’ dideoxynucleoside triphosphate substrates. When the DNA synthesis reaction is carried out in the presence of these substrate analogues, the 2’,3 dideoxynucleoside is incorporated into the growing chain but then causes termination of continued elongation because it lacks the 3’ hydroxyl on the deoxyribose at which a phosphodiester bond would form with the incoming substrate nucleotide, as illustrated in the cartoon.
In the chain termination method, four separate DNA synthesis reactions were spiked with low levels of one of the four 2’, 3’ dideoxynucleotide substrates, generating a ladder of chain termination products specific for each of the four dideoxynucleotide substrates. Thus, as in Maxam/Gilbert sequencing, the sequence of deoxynucleotides in the DNA could be read out from a ladder of DNAs of various lengths, chemically broken fragments in the case of the Maxam/Gilbert method and synthetic DNAs whose synthesis had been terminated in the case of Sanger’s 2’,3’ dideoxynucleotide method.
The chemical method of Maxam and Gilbert and the enzymatic method of Sanger can be regarded as first generation sequencing. Soon methods for automating Sanger sequencing were developed, which came to dominate DNA sequencing. In the ensuing years entirely newer, faster and cheaper methods were developed that do not rely on chain termination or gel electrophoresis. These methods have had a profound effect on the biological sciences, providing, for example, revolutionary insights into evolution, including the evolution of our own species. In the video, Gilbert describes how the world of sequencing has dramatically accelerated in the ensuing decades.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}