Chapter 9: Benzer pushes genetics to the level of nucleotides
“This great sensitivity prompts the question of how closely the attainable resolution approaches the molecular limits of the genetic material”
Recall that Schrödinger argued that the chromosome must be a “regular array of repeating units in which the individual units are not all the same.” In other words, a linear code. Once DNA was shown to be the genetic material and once its structure was solved it seemed inescapable that this linear code must reside in the sequence of nucleotides in the DNA. But genetic studies in traditional model organisms such as corn, Aspergillus, Neurospora, and the fruit fly did not offer the resolution necessary to define the boundaries of a gene or to dissect the gene at the granular level of nucleotides. This changed with the development of a powerful new genetic system by Seymour Benzer that could detect extremely rare recombination events.
Benzer (1921-2007) obtained a PhD in Physics at Purdue University in 1947. He did pioneering work in solid state physics that set the stage for the first transistors. But he became fascinated by Schrödinger's book “What is Life?”, which inspired him to switch to biology. He met Salvador Luria who suggested that he take Max Delbrűck’s phage course at Cold Spring Harbor Laboratory. He later did postdoctoral work at Caltech with Delbrűck after which he returned to Purdue where he developed a genetic system based on E. coli phage T4, the subject of this and the next chapter. In this video, Benzer describes meeting Luria and learning about Delbrűck’s course:
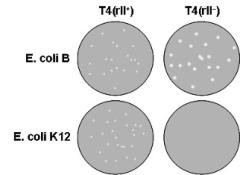
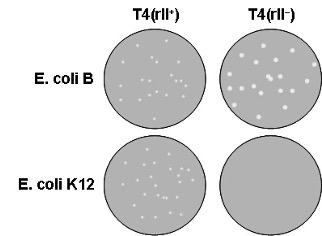
Benzer’s genetic system was based on the formation of plaques by phage T4 on a lawn of E. coli cells of a strain known as B and a second strain known as K12 (which carries a λ prophage). Mutants of T4 called rapid-lysis mutants or r mutants form plaques on strain B that are larger than the plaques formed by the wildtype phage. Genetic crosses showed that r mutations clustered in three loci called rl, rll and rlll. Importantly, and uniquely, rII mutants fail to grow on strain K12 or form plaques on a lawn of K12 cells. As summarized in the figure, rII+ (wildtype) forms plaques on both strains B and K12 whereas rII- generates oversized plaques on strain B but no plaques on strain K12.
We now know that this seemingly arcane behavior is an example of defense/counter-defense “immune” systems exhibited by many bacteria. For example, many bacteria have “restriction enzymes” that destroy phage DNA (chapter 18). And the famous CRISPR system used in gene editing evolved in bacteria as a defense against phage. In the case of K12, rII is a counter-defense to the presence of the phage λ prophage, which carries an immune gene rex that protects the lysogen against T4 that lacks the rII system.
Nonetheless, all that mattered to Benzer (or that matters to us) is simply that rII mutants grow on strain B but not on strain K12. Indeed, phage T4 growth is reduced greater than 108-fold on strain K12 when rII is mutant. Thus, restricted growth on K12 provided a powerful selection for rare recombination events between rII alleles located in close proximity to each other. In sum, Benzer could propagate rII mutants on strain B and then carry out genetic crosses between different rII alleles on K12 and do so at far greater resolution than had been possible in other genetic systems. Quoting Benzer, “This great sensitivity prompts the question of how closely the attainable resolution approaches the molecular limits of the genetic material”
What did Benzer learn from the T4 rll system? In his classic publication in 1955, Benzer made two seminal contributions. First, he pushed genetics to the level of nucleotides. Benzer knew that T4 DNA is about 2 X 105 nucleotide pairs. From measuring recombination units for various linkage groups (rI, rII, rIII, and other loci called m and tu) Benzer estimated a total of 200 recombination units across the entire T4 linkage map (one unit equals 1%). Therefore, the recombination per unit should be 10-3 (0.001) per nucleotide pair (200 units/2 x 105 nucleotides). Benzer did observe recombination frequencies as low as 0.013 but not lower. But with his extraordinarily sensitive system, he could have detected frequencies as low as 10-8. He concluded that, “Our preliminary results suggest that the chromosomal elements separable by recombination are not larger than the order of a dozen nucleotide pairs (as calculated from the smallest non-zero recombination value…).” Indeed, Benzer had pushed genetics to the level of nucleotides and concluded that the genetic material is granular. [Subsequent work has shown that the T4 chromosome is 1,500 map units long and hence Benzer’s minimal recombination unit was less than two nucleotides.] In this video, Sidney Brenner, who will take center stage in the next chapter, describes this momentous advance.
In this video, Benzer himself describes in his own words his realization that he could use the T4 rII system to carry out recombination at very high levels of resolution.
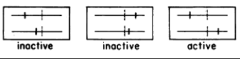
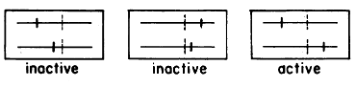
Benzer’s second seminal contribution was defining the gene as a functional unit based on a cis-trans (also known as a complementation) test, which had been developed by fly geneticist Edward Lewis. If E. coli strain K12 is mixedly infected with a T4 double mutant harboring two mutations in the rII region and wildtype T4 (the cis configuration), both mutant and wild type phage are expected to propagate with high efficiency (that is, much higher than would be expected from recombination). This is because the wildtype chromosome provides rII function for both infecting phage. On the other hand, if K12 is mixedly infected with two mutant phage each carrying a different rII mutation (the trans configuration), then two outcomes are possible. If rII consists of two functional units and if both mutations are in the same functional unit, then neither phage is expected to propagate at high efficiency. If, however, the mutations are in different functional units, then one chromosome will provide one function and the other chromosome the other function, allowing both mutant phage to propagate. In other words, the two chromosomes complement each other. These two scenarios are depicted in the diagram from Benzer’s publication in which “active” signifies phage growth and “inactive” a failure to propagate. The vertical dotted lines indicate the boundaries between the two functional units.
Indeed, Benzer observed that certain pairs of mutations were active in the trans configuration and others not. Moreover, mutations that were inactive in the trans configuration were clustered with each other in two parts of the rII region. Benzer concluded that rII consistes of two side-by-side functional units or genes, which he called A and B. He would later coin the term cistron (from cis/trans) as a definition of a functional unit or gene: “It turns out that a group of non-complementary mutants falls within a limited segment of the genetic map. Such a map segment, corresponding to a function which is unitary as defined by the cis-trans test applied to the heteorcaryon, will be referred to as a cistron.” Thus, rII consists of side-by-side, A and B cistrons.
In 1967 Benzer left Purdue and joined the faculty of CalTech where he switched fields from phage to flies. In this second phase of his career, he famously discovered genes involved in phototaxis and the circadian rhythm. Arguably, Benzer merited two Nobel Prizes. One for pushing genetics to the level of nucleotides and creating a system that would make it possible to infer that the genetic code is triplet and non-overlapping (as we come to next). And a second Prize for pioneering the discovery of genes involved in animal behavior. Sadly, he won a Nobel Prize for neither.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}