Chapter 8: Meselson and Stahl provide empirical support for semi-conservative DNA replication
“The most beautiful experiment in biology”
The double-helical model for the structure of DNA suggested a copying mechanism by which each deoxy-polynucleotide strand would serve as a template for the synthesis of a complementary copy. Such a mechanism is known as semi-conservative DNA replication in that each daughter double helix consists of an intact (conserved) parental strand and a newly synthesized, complementary copy. But in 1953 semi-conservative replication was only a model. Conceivably, for example, replication could take place by a conservative mechanism in which one daughter DNA consists of both parental strands and the other daughter DNA consists of two newly synthesized strands. In conservative replication, both parental strands would be copied into two newly synthesized strands that would pair with each other. Which mechanism was correct?
An early effort to distinguish among models for how DNA is replicated was reported by Cyrus Levinthal in 1956. Leventhal’s experiment was based on following the transfer of radioactivity from P32-labeled phage from one generation to the next. Levinthal concluded that the results were most easily understood in terms of the model proposed by Watson and Crick. Two years latter Matthew (Matt) Meselson and Frank Stahl reported on an experiment that came to be regarded as a classic in the history of molecular biology and that provided compelling evidence for semi-conservative replication.
Matt Meselson was born in 1930 and attended the University of Chicago, graduating in 1951. He spent a year at CalTech, where he took Linus Pauling’s freshman chemistry course and got to know Pauling’s children. During the summer of 1953, he was invited by Pauling’s daughter to a swimming pool party at the Pauling family home. As Meselson recalls,
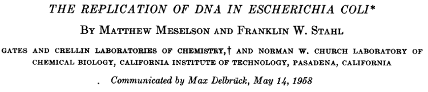{kind=link}
“… I was in the water and Pauling came up to the edge of the pool, wearing a suit and necktie. I thought, here’s the world’s greatest chemist and I’m here in the water practically naked. He looked down at me and said ‘Well now Matt, what are you going to do next year?’ I said that I was going to go back to the University of Chicago to study mathematical biophysics….Pauling looked down at me and said: ‘But Matt, that’s a lot of baloney, why don’t you come to CalTech and be my graduate student.’ I looked up at him and said: ‘OK, I will”.”’
Meselson obtained his PhD at CalTech in 1957, being Pauling’s last graduate student. He joined the faculty of Harvard University in 1960 where he has been ever since.
Frank Stahl was born in 1929 and attended Harvard College, graduating in 1951. He did his graduate work at the University of Rochester where he studied the genetics of phage T4 with Gus Doermann, a member of the Phage Group, and obtained his PhD in 1956. He met Meselson at the Marine Biological Laboratory at Woods Hole where they agreed to study DNA replication together at CalTech where Stahl did a postdoc with another phage group member Giuseppe Bertani. Stahl joined the faculty of the University of Oregon in Eugene in 1959 where he helped found the Institute of Molecular Biology.
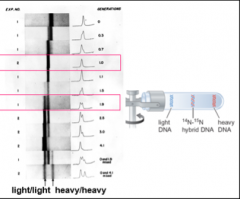
To distinguish conservative from semi-conservative modes of replication, Meselson and Stahl grew cells of E. coli on medium supplemented with the heavy nitrogen isotope 15N for many generations so that the cells were uniformly labeled. Next, the bacteria were shifted to medium containing nitrogen with mass 14N. The DNA was extracted from cells at various times after the shift and subjected to ultracentrifugation in cesium. Ultracentrifugation created a density gradient that would separate “heavy/heavy” DNA (both strands containing 15N), “light/light” DNA (both strands containing 14N), and hybrid DNA (one strand containing 14N and one 15N) (see the cartoon to the right in the figure). Since the parental DNA was from cells that had been grown on 15N, DNA extracted at the time of the shift to 14N would sediment at a heavy/heavy density. The crucial result was obtained after the cells had been shifted to and grown for one generation on 14N. In the conservative model, two species of DNA would be expected, DNA that was heavy/heavy and DNA that was light/light. This was the expectation if the parental DNA was conserved and both strands of the daughter DNA were newly synthesized after the shift to 14N-containing medium. In the semi-conservative model, the duplex would unwind and each strand of the parental DNA would serve as a template for the synthesis of a complementary, 14N-containing strand. If this were the case, then only a single density band should have been seen, corresponding to a heavy/light, intermediate density.
The experimental findings of the historic publication are shown to the left in the figure with results obtained after one generation (1.0) after the shift highlighted with a red box. Also, highlighted are the results seen after approximately two (1.9) generations. Labeled at the bottom of the figure are the densities for light/light and for heavy/heavy duplexes. It can be seen that all of the DNA shifted to the intermediate, heavy/light density after one generation, consistent with the semi-conservative model. Moreover, after a further generation, two species were seen, heavy/light and light/light, also in full agreement with the semi-conservative model in that after the second round of replication, the light strand of the hybrid DNA would serve as a template for a light strand, resulting in fully light duplex. Meanwhile, the heavy strand of the hybrid would serve as a template for a newly synthesized light strand, resulting once again in a hybrid duplex.
In a letter to Watson shortly after completing the experiment (November 8, 1957), Meseslson describes the results as "Clean as a whistle! Who would have imagined that..." Indeed.
Did the Meselson-Stahl experiment truly prove that replication was semi-conservative? Australian scientist John Cairns, who would later become the Director of the Cold Spring Harbor Laboratory, raised the esoteric possibility in a 1962 letter to Nature that perhaps DNA consists of a pair of double helices. In such a case, no unwinding would be necessary to obtain the results seen in the Meselson-Stahl experiment. Rather, each member of the pair would generate a daughter that was light/light (as in the conservative model), resulting in a pair of double helices in which one duplex was heavy/heavy and one was light/light. In this scenario the overall density after one generation would be hybrid even though replication was conservative! Cairns, however, put this unlikely model to rest by measuring the length of a phage DNA (lambda) and determining its mass, enabling him to conclude that DNA does indeed consist of only a single double helix.
Cairns was a great admirer of the Meselson-Stahl experiment and having proven beyond doubt that DNA replication must be semi-conservative, he famously referred to it as “The most beautiful experiment in biology” (as quoted by Holmes in Meselson, Stahl and the Replication of DNA). Cairns apparently later referred to Mendel’s experiments as the most beautiful in biology in an exchange with Stahl that is amusingly recounted by Stahl in the iBiology video.
{kind=link}
Another pioneer who helped validate the Watson-Crick model for base pairing and who opened a new era in the toolkit available to molecular biologists was Paul Doty (1920-2011). Doty was noted both for his contributions to molecular biology and to nuclear arms control. He and his co-workers discovered that the two strands of DNA could be separated and then correctly re-annealed to re-form a double helix. He recruited Jim Watson and Matt Meselson to Harvard and founded the Department of Biochemistry and Molecular Biology. In the arena of nuclear arms control, he went to the Pugwash nuclear arms control meeting in 1955 in follow up to an appeal by Bertrand Russell and Albert Einstein. He then brought Russian and American scientists together in pursuit of peace, making more than 40 trips to the Soviet Union. He later founded the Belfer Center of Science and International Affairs at Harvard’s Kennedy School. His wife, Helga Doty, made fundamental contributions to our understanding of the collagen component of connective tissue. An amusing anecdote about Doty (Candid Science II by I. Hargittai) highlighting that the genetic material had at last become part of common folklore is as follows: “…some time in the early sixties Paul Doty….saw displayed on a New York paper stall lapel buttons that bore the legend ‘DNA’. Supposing that the letters must stand for some political slogan also (Democracy not Anarchy?), he asked the paper-seller what they signified. ‘Get with it, mac’, came the reply, ‘dat’s de gene’.”
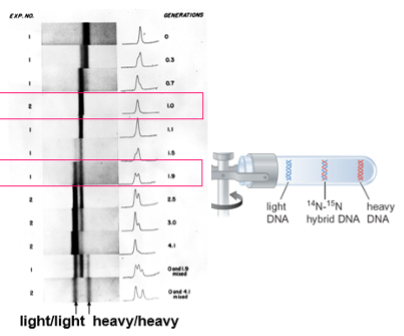
In two classic back-to-back publications in the Proceedings of the National Academy of Sciences (1960), Doty and his team demonstrated that the strands of DNA could be separated by heating and allowed to correctly re-anneal by gradually lowering the temperature. This was demonstrated by two independent methods. In one, the ability of “transforming principle” to convert pneumococcus to streptomycin resistance [as pioneered by Harriet Ephrussi-Taylor in Avery’s laboratory (chapter 3)] was shown to be destroyed by incubation at 100oC but substantially restored by gradually lowering the temperature (left-hand figure below). The second method took advantage of the observation that denatured DNA has a greater absorbance of ultraviolet light at 260 nm than native double-stranded DNA. Thus, native DNA exhibits an abrupt increase in absorbance when increasing temperature reaches 80 oC to 100 oC (the DNA melting temperature) (right-hand figure). The increase in absorbance can be partially reversed by cooling the DNA. This partially renatured DNA is once again sensitive to increasing temperature exhibiting a marked increase in absorbance above 80 oC.
The discovery that DNA could reanneal and, as was later shown, could hybridize with RNA of complementary sequence ushered in an era of powerful technical advances that greatly accelerated advances in molecular biology. These advances include the capacity to detect specific messenger RNAs and specific genes and the capacity to amplify specific DNA sequences by the Polymerase Chain Reaction (chapter 21).
Finally, we digress to highlight a common oversimplification about the basis for the stability of the double helix in an aqueous environment, namely, that the strands of DNA are held together by hydrogen bonds. This is misleading. Hydrogen bonding between A and T and between G and C is not a major reason double-stranded DNA is more stable than single stranded DNA. This is because the hydrogen bonds between the bases largely replace hydrogen bonds to water in single-stranded DNA. What then is the source of energy to stabilize the double helix? One source is the increase in entropy when ordered water molecules assembled around the hydrophobic surfaces of the bases in single-stranded DNA are released when the double helix is formed (known as “the hydrophobic effect”). Also contributing to the stability of the double helix are π-π interactions between the electron clouds of the aromatic rings of the stacked bases. Interestingly, G:C-rich DNA is more stable (has a higher melting temperature) than A:T- rich DNA. But this is not principally due to G:C having three hydrogen bonds and A:T two. Rather, it is because stacking interactions with G:C base pairs are more favorable than stacking interactions with A:T base pairs.
We close by noting how far our understanding of the genetic material progressed in less than two decades: from the early 1940s when it was discovered that DNA and not protein is the stuff of genes to the late 1950s when the structure of DNA was solved, revealing the basis for how the genetic material is duplicated and setting the stage for deciphering the linear code of chromosomes.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}