Chapter 13: Gilbert, Műller-Hill and Ptashne Isolate Repressors and Demonstrate That They Bind to DNA
“An isolated repressor is shown here to bind specifically and with high affinity to DNA, strongly suggesting that, in vivo, repressors block the transcription from DNA to RNA by binding directly to the DNA.”
Jacob and Monod provided compelling genetic evidence that the lac operon and phage λ genes are subject to negative regulation by repressors. But they left two major questions unanswered: what is the chemical nature of these repressors and how do they work? These questions were answered within the next decade by Wally Gilbert and Mark Ptashne, who were in an intense competition with each other in the Harvard Biological Laboratories.
Wally Gilbert was born in 1932. He trained as a physicist and joined the Physics Department at Harvard University as an assistant professor in 1959. Gilbert became friends with Jim Watson, leading him to become interested in molecular biology and move to the third floor of the Biological Laboratories and run a joint laboratory in the 1960s. He eventually left the Physics Department to join Watson’s Department of Biochemistry and Molecular Biology. During this period Gilbert contributed to the discovery of messenger RNA and the isolation of the Lac repressor and the demonstration that it binds to DNA. He then went on to invent a method to sequence DNA (chapter 20), for which he shared a Nobel Prize with in 1980 with Fred Sanger (and also Paul Berg for recombinant DNA; chapter 19). With Nobel Laureate Phil Sharp and others he founded one of the first biotech companies, Biogen, and left Harvard to serve as CEO. He returned to Harvard in 1985, serving as Chairman of the Harvard Society of Fellows. He retired from Harvard in 2001, launching an artistic career in digital photography.
Gilbert’s quest to isolate the Lac Repressor was carried out with German scientist Benno Műller-Hill. Their strategy was to purify a protein that would bind the artificial Lac inducer IPTG, which had been used by Jacob and Mond. They used a dialysis assay in which binding was monitored by increased concentration (excess binding) of radioactive IPTG (C14-IPTG) inside a dialysis bag as compared to outside the bag.
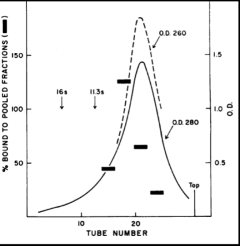
Partially purified repressor was subjected to centrifugation through a gradient of glycerol. Pooled fractions from the gradient were then tested for the ability to bind C14-IPTG by the dialysis assay. The figure shows the profile of such a gradient in which the top of the gradient is on the right and separation according to increasing size proceeds from right to left. Samples from the gradient were collected in tubes as indicated by the horizontal axis. The solid curve shows the distribution of protein as judged by UV absorbance at 280 nm. The thick, horizontal bars show the results of the inducer binding assay. It can be seen that a peak of C14-IPTG-binding activity sediments at a position slightly larger than the bulk of the protein. Thus, as judged by this purification step the repressor sedimented as a relatively homogenous species.
Gilbert and Műller-Hill describe the properties of the repressor as follows: “The ability to bind IPTG is not attacked by RNase or DNase. It is destroyed by pronase. The binding site can be inactivated by temperatures above 50oC.” Based on this evidence they conclude that the repressor is a protein, addressing the first of Jacob and Monod’s two questions. They tackled the second question by preparing radioactive repressor and asking whether it would bind to lac DNA.
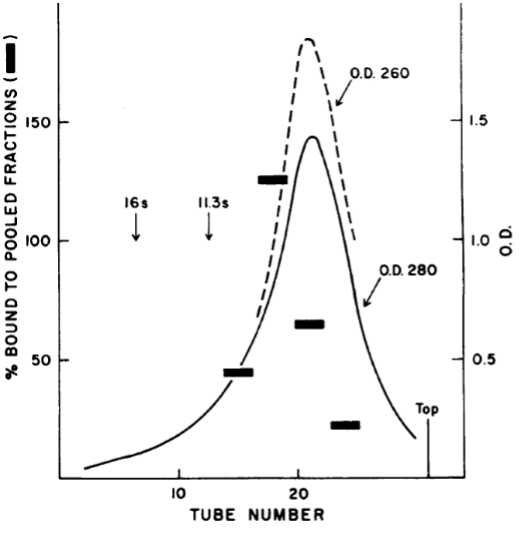
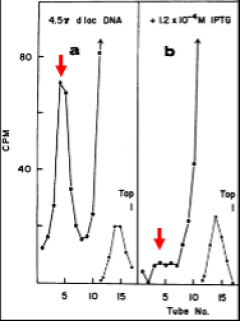
Radioactive repressor was sedimented through a glycerol gradient in the presence of lac-containing DNA. As seen in panel a (left), radioactivity sedimented at a position corresponding to tubes 4 and 5 as highlighted with the red arrow. Panel b (right) shows that little radioactivity sedimented at this position when centrifugation was carried out in the presence of IPTG. Evidently, then, repressor binds to lac DNA and this binding is prevented by the presence of inducer. Moreover, an additional experiment showed that binding to lac DNA was specific as no binding was observed with DNA not containing lac DNA.
Finally, certain mutants of lac known as Oc cause constitutive synthesis of β-galactosidase. Oc mutants are cis-acting as constitutive synthesis is observed in the presence of a wildtype copy of lac. Using their binding assay based, Gilbert and Műller-Hill found that binding of repressor to lac DNA containing the Oc mutation was conspicuously weaker than binding to wildtype lac DNA. Thus, in answer to the second of Jacob and Monod’s questions, the repressor evidently acts by binding to the lac operator and does so in a manner that is blocked by the presence of inducer.
All this was carried out in hot competition with a second investigator in the Biological Laboratories, Mark Ptashne. Ptashne, who was born in 1940, earned his PhD in 1968 with Matt Meselson in the Harvard Biological Laboratories. He was appointed a Junior Fellow of the Harvard Society of Fellows (1965-1968) after which he became a Lecturer and was promptly promoted to Professor in 1971. Ptashne is known for the first demonstration of specific binding between a protein (λ repressor) and DNA. His book, A Genetic Switch, on the regulatory circuit governing the switch between the prophage and lytic growth states of λ is a classic in the field of gene control. Ptashne went on to study gene control in yeast. He founded (like Gilbert) one of the first biotech companies, The Genetics Institute, in 1980. He won the Lasker Award in 1997. Ptashne is also a violinist who would perform at meetings at the Cold Spring Harbor Laboratory. He eventually left Harvard and is now at the Memorial Sloan-Kettering Cancer Institute.
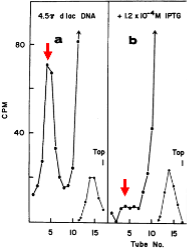
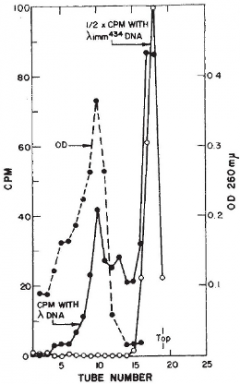
Ptashne isolated the phage λ repressor by comparing the radioactive profile of a mixture of H3-labeled protein from cells infected with wildtype phage and C14-labeled protein from cells infected with λ mutant for the phage repressor gene (C1). Chromatography revealed a protein that was uniquely labeled with H3. The Ptashne paper was published a few months after that of Gilbert and Műller-Hill. But since the Ptashne isolation of repressor was based on radioactive labeling of the protein, he was immediately prepared to ask whether repressor acts by binding to DNA. Accordingly, he subjected radioactive repressor to sedimentation through a sucrose gradient in the presence of phage λ DNA and DNA from a closely related λ phage (λ434) that is not subject to repression by the λ repressor. (λ434 has its own repressor and operator and is said to have a different “immunity” than that of λ.) The striking result (figure to the left) was that radioactive λ repressor exhibited a fast-sedimenting peak (tube 10; solid line with filled circles) when the experiment was carried out with λ DNA but not when it was carried out with λ434 DNA (tubes near the top; solid line with open circles).
Because Ptashne already had radioactively-labeled repressor, he was able to carry out his DNA-binding experiments more quickly than Gilbert and Műller-Hill. So whereas the latter won the race to isolate a repressor, Ptashne was able to complete and publish his demonstration of specific binding to λ DNA a few months earlier than Gilbert and Műller-Hill. This was a stunning race representing a major advance in molecular biology, an advance that provided a brilliant capstone to the pioneering work of Jacob and Monod in laying the groundwork for the concept of negative control and repression. The video is an excerpt from the TV documentary “The Scientist,” which captures the rivalry in the race for the repressor.
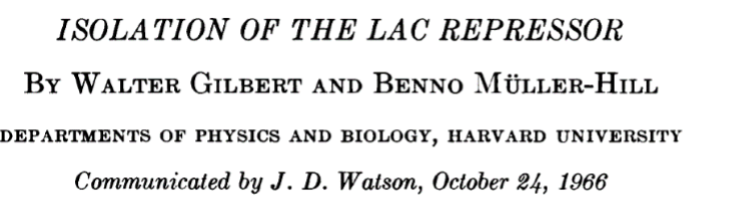{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
The race for the repressor was no doubt an important chapter in molecular biology, but the TV documentary (video) went a bit over the top in claiming that “when the battle is won it will be more important than the atomic bomb.”
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}